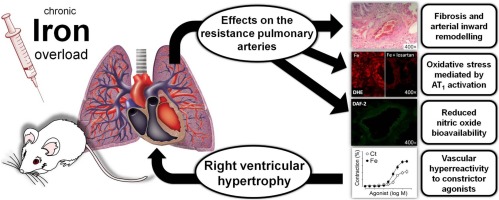
“ferric ammonium citrate-induced interleukin-1b production is dependent on caspase-1 and is significantly inhibited by an Fe2+-specific chelator. Ferric ammonium citrate consistently induced interleukin-1b secretion in THP1 cells, but not in NLRP3-deficient THP1 cells, indicating a requirement for the NLRP3 inflammasome. Additionally, activation of the inflammasome is mediated by potas- sium efflux, reactive oxygen species-mediated mitochondrial dysfunction, and lysosomal membrane permeabilization. Thus, these results suggest that monocytes/macrophages not only sequestrate iron during inflammation, but also mediate inflammation in response to cellular labile iron, which provides novel insights into the role of iron in chronic inflammation.”
-
By using this site you agree to the terms, rules, and privacy policy.
-
Charlie's Restoration Giveaway #2 (Entire Home EMF Mitigation & Protection Along With Personal Protection) - Click Here To Enter
-
Dear Carnivore Dieters, A Muscle Meat Only Diet is Extremely Healing Because it is a Low "vitamin A" Diet. This is Why it Works so Well...
Rest the rest of this post by clicking here
-
The Forum is transitioning to a subscription-based membership model - Click Here To Read
Click Here if you want to upgrade your account
If you were able to post but cannot do so now, send an email to admin at raypeatforum dot com and include your username and we will fix that right up for you.
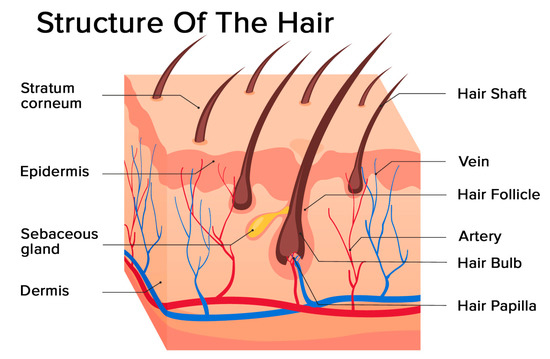
Activation of the NLRP3 inflammasome by cellular labile iron
- Thread starter Jamsey
- Start date








EMF Mitigation - Flush Niacin - Big 5 Minerals