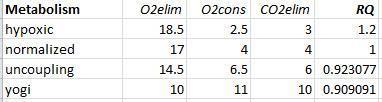
Interesting. An IC would definitely not suit your purpose. Thanks for explaining what you are working toward in more detail.visionofstrength said:Yes, good example. Those directly measure CO2 and O2 but display the conventional "caloric" calculation, as described in the standard theory of RQ.Blossom said:I wonder if an indirect calorimeter could help?
In my view, if Peat is right about hypoxia and uncoupling, the calorimeters are not calculating the "calories" the way they think, because the RQ standard theory ignores the effects of hypoxia or uncoupling (which don't follow the RQ standard theory about calories).
-
By using this site you agree to the terms, rules, and privacy policy.
-
Charlie's Restoration Giveaway #2 (Entire Home EMF Mitigation & Protection Along With Personal Protection) - Click Here To Enter
-
Dear Carnivore Dieters, A Muscle Meat Only Diet is Extremely Healing Because it is a Low "vitamin A" Diet. This is Why it Works so Well...
Rest the rest of this post by clicking here
-
The Forum is transitioning to a subscription-based membership model - Click Here To Read
Click Here if you want to upgrade your account
If you were able to post but cannot do so now, send an email to admin at raypeatforum dot com and include your username and we will fix that right up for you.
Optimal Diet For Increasing Lifespan
- Thread starter haidut
- Start date
EMF Mitigation - Flush Niacin - Big 5 Minerals